Franco Manni
Cultural Traumas, Intellectual Fallacies and Moral Mistakes. The current debate about the existence of human races

"Race" is synonymous with "subspecies", that is, a sub-set of whatever species of living beings.
If there is no interfertility between one species and another species, there is indeed interfertility between a race (subspecies) and another race, as we have seen for centuries in the equine, canine and even human races.
The divisions in groups of humanity are evident based on religious beliefs (Islam, Buddhism, etc.), with respect to the nationality i.e. language (Francophones, Arabs, Anglo-Saxons, Hispanic, etc.). And a division with respect to
biological characteristics is also evident (black, Asians, mulattos, whites, etc.) These biological characteristics are precisely the several subspecies or races of our one species homo sapiens ...But because of the tragedy of Nazism and the Holocaust, and because of racism, even later and now, there is a moral and intellectual trauma in the Western conscience and this trauma causes the political ideology to confuse racism, which is a political fact, with the existence of races, which is only a biological fact.
Racism means to believe that one race is superior to the others and also that, because of this presumed superiority, it can afford to take off others’ rights and limit or suppress them in order to dominate or persecute the allegedly ‘inferior’ races.
‘Race’ means that a
group of individuals can be grouped together by somatic similarities that unite them more than with other individuals.What features? They are not those of the internal organs such as pancreas or blood composition, etc., but the external, visible, which strike at first sight: skin colour, shape of the nose, glabrity or skinniness of the skin etc.
Those who deny the existence of races (and therefore of these different physical criteria of grouping), certainly will never be able to convince ordinary people in whose eyes these differences are evident and will never disappear. Or to convince the police or the employers!
But they commit the grave error both intellectual and moral to give this message to people: " in order to affirm that all human beings have equal dignity and therefore must have equal rights, we must affirm that they are equal in their material and
de facto situations , like, for example the physical characteristics".... but this is a serious mistake, because equal dignity must be recognized not if we are equal (all Christians, all heterosexuals, all healthy, all fascists or all communists for that matter, all of the same biological group)! NO!
Equal dignity must be recognized precisely when we see that materially, economically, linguistically by political opinion, by religious faith, by somatic characteristics, by sexual orientation WE ARE DIFFERENT. Equal moral and legal dignity, despite the material and social de facto differences!!!
Otherwise the same concept of tolerance disappears
. In fact, it is absurd to say that you practice the virtue of tolerance towards those who are equal to you by religion, social class, nationality, etc.!Tolerance is practiced precisely towards those who are different from you!
If you really do not want to hit the sick hypersensitivity of these people who deny the existence of the races, you should also use another word (biological groups, subspecies, genetic classes ... you choose!)
... but it is an intellectual and moral error to deny reality!
I enclose a few documents on the existence of the races:
-
the first is the entry "human races" from the Encyclopaedia Britannica (1997)
-
the article of A.W.F. Edwards, ‘Human Genetic Diversity: Lewontin’s Fallacy’, BioEssays 25:798–801, 2003 Wiley Periodicals, Inc.; where the author traces the current academic fashion back to biologist R.C. Lewontin’s essay of 1972
-
a statement by Richard Dawkins, who, in his book The Ancestors’ Tale (2005), explains why Lewontin’s argument is false.
-
a chart from the book by SJ Gould (professor of biology at Harvard and probably the greatest scholar of Darwin of the twentieth century) that cites a study by Luca Cavalli Sforza (probably one of the most important living geneticists) and reproduces one of his diagrams that distinguishes the biological subgroups of our human species (the human races!) according to the quantity of different chromosomes.
-
The poster made by Prostate Cancer UK to warn black men that the probability they get a prostate is double than that one of the other men.
-
The program of the conference "Embodiment and Race", held at Clemson University (North Carolina) on 11-13 April 2019;
-
The paper I delivered at that conference.
© Encyclopedia Britannica – 1997
GENETICS OF RACES AND SPECIES DIFFERENCES
Arthur
Robinson, M.D. Professor of
Biochemistry, Biophysics, and Genetics and of Pediatrics, University of
Colorado, Denver. Senior staff member, National Jewish Center for Immunology
and Respiratory Medicine, Denver.
Francisco
Jose Ayala. Donald Bren Professor of
Biological Sciences, University of California, Irvine. Author
of Evolving: The Theory and Processes of Organic Evolution and others.
Russell
Howard Tuttle. Professor of
Anthropology, University of Chicago. Author of Apes of the World: Their
Social Behavior, Communication, Mentality and Ecology.
The
nature and origin of R A C E S
and species are dealt with in
the article EVOLUTION, THE THEORY OF, particularly in the section
Species and speciation. Here it is necessary to consider only the genetic
composition of the R A C E and species differences in sexually
reproducing and outbreeding organisms.
In
general, species are considered to be populations of organisms between
which breeding is impossible or significantly limited under natural
conditions. Subgroups of an individual species with distinctive
phenotypes form R A C E S, members of which can interbreed with members of other
R A C E S of that species. The geneticist Curt Stern defined a R A C E as a
group more or less isolated geographically or culturally who share a
common gene pool and who, statistically, are somewhat different at some loci
from other populations.
In
naturally occurring populations a species may split into R A C E S because
of a gradual geographical separation and eventual rift between formerly
interbreeding groups. Such R A C E S, which inhabit different territories,
are called allopatric. If these R
A C E S are brought together, they assume a
sympatric status, interbreed, exchange genes, and fuse into a single
genetically variable population. R A C E S of human beings and of
certain parasites are exceptional because they can coexist, at least for a time,
sympatrically. In humans, social rather than geographical or biological
factors slow down interbreeding and R A C E fusion. Distinct species
may, on the other hand, be either allopatric or sympatric. The exchange of
genes between species populations is prevented not only by geographic
distance (as with R A C E S) but also by genetically based reproductive
isolating mechanisms. Reproductive
isolation is achieved by a variety of means: differences in preferred
habitats, in breeding seasons, in sexual attraction and courtship rites,
in sexual structures (flowers in plants and genitalia in animals);
incompatibility of sex cells; inviability of the hybrid progenies;
sterility of the hybrids; and weakness of the gene recombination products of
the gene complements of the species.
R A C E differences are more often quantitative than qualitative; racially distinct populations of a species differ usually in the frequency of certain genes rather than the presence or absence of certain genes.
Studies
on human blood groups have revealed
some instructive situations. As discussed earlier, the four "classical"
blood groups O, A, B, and AB are due to three alleles of a gene. Most
human populations have individuals of all four types, and even parents and
children, as well as brothers and sisters, may belong to different
blood groups. However, some blood groups, and hence the gene alleles that
produce them, are more frequent in some countries than in others. The
gene for A blood, for example, increases in frequency from east to west
in Europe; B blood is most frequent in some populations of India, Tibet,
Mongolia, and Siberia. A majority of American Indians apparently had,
before the arrival of Europeans and Africans, the O blood group only;
however, the tribes of the Blackfoot
and Bloods had the highest known frequencies of A blood, which is also very
frequent among the Lapps in northern Europe (see also BLOOD: Blood
groups). Human R A C E S differ certainly in many genetic
traits, not in blood groups alone. Some theorists, imagining the human population
as it might have been about 4000 BC, have speculated that there were perhaps
five major R A C E S, one for each inhabited major landmass.
The
major human R A C E S are separated by their most readily recognized
characteristics, such as skin colour, body size, and facial morphology.
In modern times many of the barriers, both geographic and cultural, between the
R A C E S have weakened.
Since most of the external differences between R A C E S are polygenically
and environmentally determined, interracial matings produce offspring
that, in general, have a phenotype intermediate between those of the parents.
Of the total number of loci existing in the human
species, numbering at least 100,000, the majority of their alleles probably
are present among the members of each R A C E.
Although
one population of alleles--for example, those for dark skin colour--may be
common in one R A C E and rare in another, each particular skin-colour
allele occurs in both R A C E S. In fact, studies involving many
polymorphic loci (loci with two or more alleles occurring with a frequency
of at least 1 percent) have revealed that the allelic diversity among
individuals within a single R A C E is greater than that between R A C E S.
Intelligence is a highly variable characteristic that some have
claimed varies among different human R A C E S. This characteristic is
difficult to define and even more difficult to measure. In addition, it is
significantly influenced by environmental factors. Finally, the trait
certainly varies more within members of a R A C E than between R A C E S. R
A C E S are genetically open systems, and gene exchange between R A C E S
does take place. Species, by contrast, are genetically closed systems
in which gene exchange is rare or absent. R A C E differentiation is
reversible; hybridization or intermarriage may cause R A C E S to merge
into a single population. It is an error to think that in a population
resulting from R A C E hybridization all individuals will be alike; in point of
fact, such hybrid populations show a remarkable diversity of individuals.
Species differentiation is irreversible.
R
A C E S are populations, and an individual may have a genetic endowment that
can occur in two or more different R A C E S or that is not common in
any R A C E. An individual belongs, however, to only one species, unless
that individual is a species hybrid.
Mules, hybrids between the horse and donkey, are sterile because of
abnormalities in the processes of sex-cell formation in their gonads.
Sterility of hybrids between species, if viable hybrids between them
can be obtained at all, is observed very frequently, though some
experimentally obtained species hybrids have proved to be fertile. Scientists
have asked what causes the development of the gene-frequency differences
between populations that live in different territories, or, in other
words, what makes these populations racially distinct.
The probable explanation is that genetic differences
between populations arise in most cases through natural selection in
response to the local environments that prevail in the territories they
inhabit. It is, however, very difficult to verify this explanation in many
concrete instances of R A C E differentiation. For example, it is
probable that the dark skin pigmentation of many human populations that
live, or have until recently lived, in tropical and subtropical
countries protects them from sunburns. It is probable also that the light
skin colours of the natives of Europe facilitate the acquisition of
vitamin D in regions with deficient sunshine.
The
evidence for even these hypotheses is not as conclusive as might be
desired. But when it comes to such racial traits as hair form and shapes of
the nose, of the lips, and of the cheekbones, no acceptable evidence of
adaptive significance is available. The situation is no better with R A
C E S of animals and plants: for most racial differences the adaptive
significance is unknown. Attempts have been made to envisage
factors other than natural selection that could be responsible for genetic
differentiation of populations. Appeals are frequently made to
pleiotropism of the gene action; a visible R A C E difference may in itself
be neither adaptive nor unadaptive, yet it may be only an outward sign
of an underlying physiological difference that is adaptively important.
An elegant example is the coloration of onions--red and purple bulbs are
resistant to the attacks of a smudge fungus, while white bulbs are
highly susceptible. A R A C E trait may also be important as a sexual
recognition mark, or it may play a role in the courtship ritual. A
most interesting possibility that should be seriously considered is that
some differences between populations may be due to random
genetic drift. As was discussed above, genetic drift acts on small
populations. Suppose that a species lives in many
isolated colonies, some of them consisting of only tens or perhaps
hundreds of individuals. Chance events may cause the gene frequencies
in the different colonies to drift apart. How important this random genetic
drift may be in R A C E differentiation is controversial. That genetic
drift does occur is certain; a simple example is that in small villages a
sizable fraction of the inhabitants sometimes have the same surname,
and different surnames are frequent in different villages. Increasing
or decreasing frequencies of the surnames evidently go together with
increases or decreases of the frequencies of certain genes that the
ancestors of the people with these surnames carried. As discussed, genetic
drift can also arise from the founder effect. When the populations of
new colonies founded by a small number of individuals expand, they will be
found to differ genetically from each other and from the ancestral
population. Natural selection will then come into operation, giving rise to
new balanced gene pools. The founder
effect was probably important in the development of some human populations.
Many tribes and local R A C E S may be the descendants of small numbers of
original migrants and settlers. Whether the random genetic drift alone
can explain the origin of the gene complexes that differentiate R A C E S or
species is very doubtful. The point is, however, that genetic drift and
natural selection are not mutually exclusive alternatives; it is not one or
the other but the interaction of both that brings about R A C E
differentiation. The founder effect is a special case of random genetic
drift. The gene pool of a colony derived from a single immigrant or
several pairs of immigrants may need a restructuring by natural selection
to become properly adapted to the new environment.
Human
Evolution R A C E AND POPULATION Definitions and
terminology.
The
term R A C E as applied to humans has been variously used--by politicians,
military leaders, philologists, human biologists, demographers, and
historians. Some "R A C E
S" constitute language groups, often of peoples whose only kinship is
that they speak a common language. Such was the original meaning of the
so-called Aryan R A C E. Some "R A C E S"
are simply hypothetical, invented to embrace present distributions of
such genetic (hereditary) characteristics as stature or hair colour--e.g.,
the Nordics. (The word Nordic also has been given a political meaning,
referring, despite their differences in physical characteristics, to
peoples in northern Europe.)
R
A C E has been variously applied to national or cultural groupings, as in
the days when English writers referred to an Irish R A C E and to a
Scottish R A C E. As used in census and other applications, the designation R A C E
often groups different peoples for administrative convenience; thus, the
category Hispanic may group people from Meso-America, the Caribbean,
South America, and the Philippines who may differ considerably in their
racial origins. "R A C E" also has been applied to
human groups inferred to have existed on the basis of archaeological
discoveries; the Etruscan R A C E is an example. Various religious
groups who may or may not have common ancestry sometimes are called R A
C E S--the Jewish R A C E, for example. By extension of biblical thinking
and in honour of Shem, son of Noah, a Semitic R
A C E was conceived in an effort to describe peoples who spoke Semitic
tongues, some of whom may have learned their language more recently
than others.
All
of those uses of the term R A C E are separate and distinct from its
biological meaning in classification (taxonomy)--the natural
divisions or groupings below the species level. As such, R A C E differs from breed or line,
which refer to artificially established groups maintained by intensive
selection or by deliberate hybridization. Just as the term R A C E is often
too broadly applied to the entire species of man (as in the human R A C
E), particular R A C E names invented to explain distributions of observable
physical characteristics of human populations are not biologically
meaningful.
The
misuse of the word R A C E--particularly the manner in which it was employed
by Nazi Germany--had led workers to search for alternate terms. Some
biological descriptions refer to human stocks, one intention for this
being to avoid political overtones. Other writers have favoured the
word division in lieu of R A C E, again apparently to escape what may be
perceived as offensive connotations. Other references to these human
groupings include strain (without implying the equivalent of purebred strains
of laboratory animals); variety (although the specific botanical
meaning does not apply to human R A C E S as ordinarily constituted);
and ethnic group, which, although generally meaning cultural or
political groupings (e.g., Macedonians, Croats, Magyars, or Slovenes),
is at times used with exactly the same biological meaning as R A C E.
With
the advent of population genetics, establishing gene frequencies in
specific populations, many
workers have come to prefer the word population for taxonomic
purposes. Populations so defined, however--such as San (Bushman), Ainu, Lapp,
Eskimo, Coloured (South Africa), or Micronesian--are often the same
groupings that have been or can be called R A C E S. Still, population is a
useful addition for such linguistically and genetically distinct groups
as the Basques and is an easier concept to explain.
The
term geographic, or continental, R A C E is often used to describe
populations that occupy a broad geographic range. Likewise, local R A C
E is used for populations in a more restricted area, and microR A C E may
correspond to a single, extended breeding population. These natural
groupings, which reflect geographic (and therefore reproductive) isolation,
display a range of genetic differences that are the focus of much
research. The ultimate questions are how long the R A C E S (or populations)
have been distinguishable and what processes brought about the
distinctions. What the different geographic R A C E S are called
is to some extent unimportant as long as the same terminology is employed by all;
such traditional designations as white, yellow, and black, however, are
clearly inappropriate. The designations for local R A C E S and microR
A C E S are similarly unimportant, except for communication and for the
sensitivities of the people themselves. It has long been a practice on
the part of some human taxonomists to convert place-names into taxonomic
names by adding the suffix -id (e.g., Pennsylvanid, Montanid) or the
suffix -oid (Capoid for the Cape peoples of South Africa). Geographic terms, without
suffixes, also suffice (hence, the Mediterranean R A C E) or are used in
conjunction with language groupings, where justified (e.g.,
Azteco-Tanoan). Often reference is made to particular national or cultural
groupings, such as Finns, because available data are so arranged, or to
artificial groupings (e.g., Ghanaians or Vietnamese) until further
information can establish more precisely the makeup of those groups.
Even a designation as being from a city may not be enough, given demographic and
genetic differentiation within cities, Tokyo being a typical example.
Human
Evolution Geographic R A C E S.
Naturally occurring (i.e., produced by natural,
usually geographic separation
of human groups) R A C E S of the human species are by no means
identical in number of members or degree of genetic differentiation. There
are small groupings of a few hundred to a few thousand individuals,
some slightly and only recently isolated reproductively from adjacent
people. Other equally small groups may have been mating apart from the
rest of mankind for centuries or even for thousands of years. Members
of some human R A C E S number in the hundreds of millions (as the peoples
of modern Europe) or in the billions (as in Asia).
It
is both useful and meaningful to identify the very large human groupings
that often correspond to continents or other major geographic areas as
geographic R A C E S, a term extensively used with other life forms.
Geographic R A C E S are numerically large, containing within them
smaller groups of reproductive isolates (breeding populations). The reasons
for the large groups' geographic delineation are usually clear. The
Indians of the Americas were
reproductively separated from the peoples of other geographic regions
for many thousands of years. Thus, they have come to differ genetically from
the rest of mankind and even from those Asians from whom they stemmed.
The Australian Aborigines
similarly constitute a geographically defined group of local R A C E S
(see below) separated for millennia from the rest of the world, except for
some slight contact, until the late 18th century. Human
Evolution The antiquity of Homo sapiens. It is a
curious fact that, although evidence for the evolution of man is extensive,
direct fossil evidence of the earliest members of the species Homo
sapiens is relatively scarce. The species H. sapiens (of which the modern
human R A C E S comprise a number of different geographic varieties)
may be defined in terms of the anatomic characters shared by its members.
The definition for prehistoric representatives of the species must be
limited to skeletal characters, the only remains to be found, and includes
such features as a mean cranial capacity of about 1,350 cubic centimetres
(82 cubic inches), an approximately vertical forehead, a rounded
occipital (back) part of the skull with a relatively small area for the
attachment of the neck musculature, jaws and teeth of reduced size,
small canine teeth of spatulate form, the presence of a pointed or
projecting chin, and limb bones adapted to a fully erect posture and
gait. Any skeletal remains that conform to this pattern to an extent that
precludes classification in other groups of higher primates must be
assumed to belong to anatomically modern H. sapiens. In the past there
was a tendency to create entirely new species of Homo on the basis of
fragments of prehistoric human skeletons, even though the remains
showed no significant differences from modern man. This tendency was
prompted by the supposed antiquity of the remains or by a failure to
realize how variable some features are even in modern man. The species of
the genus Homo that immediately preceded H. sapiens was H. erectus, and
it is most likely that sapient humans (H. sapiens) evolved from H.
erectus.
A. W. F. Edwards, Human Genetic Diversity: Lewontin's fallacy
Richard Dawkins, in his book The Ancestors’ Tale (2005), explains why Lewontin’s argument is false:
"
It is genuinely true that, if you measure the total variation in the human species and then partition it into a between-race component and a within-race component, the between-race component is a very small fraction of the total. Most of the variation among humans can be found within races as well as between them. Only a small admixture of extra variation distinguishes races from each other. That is all correct. What is not correct is the inference that race is therefore a meaningless concept. Lewontin’s view of race has become near-universal orthodoxy in scientific circles.We can all happily agree that human racial classification is of no social value and is positively destructive of social and human relations. That is one reason why I object to ticking boxes on forms and why I object to positive discrimination in job selection. But that doesn’t mean that race is of "virtually no genetic or taxonomic significance." This is Edwards’s point, and he reasons as follows. However small the racial partition of total variation may be, if such racial characteristics as there are highly correlated with other racial characteristics, they are by definition informative, and therefore of taxonomic significance.
"

Birmingham, both of which have high populations of black men.
The awareness drive is focusing on black men because of the increased risk of developing the disease- on average one in four black men in the UK will get prostate cancer at some point in their lives, as opposed to other men who have a one in eight chance of developing it.
Embodiment and Race Conference
Clemson University, April 11-13, 2019
THURSDAY April 11th
All Thursday sessions, apart from the keynote address at 5:30pm, will all be Bowfin (Outdoor lab). The keynote address is going to take place at McKissick Theatre, 720 McMillan Rd, Clemson, SC 29631
8:30-9:30 Yoga and Meditation, Renee Gahan
8:00 – 9:30 Breakfast
9:30 -11:00 Registration: Outdoor Lab
11:00-11:15 Opening Remarks: Edyta Kuzian, Conference Director
- Session 1: Concepts of Race and Racism Moderator:
- Session 2: Race and Embodiment Moderator:
- Session 3: Decolonization and Race
- 5:30-7:00 KEYNOTE: Alia Al-Saji, McGill University "Decolonizing Phenomenology: Merleau- Ponty, ‘Primitive’ Peoples, and Prepersonal Life"
- at McKissick Theatre
- Session 4: Embodied Experience and the Political Moderator:
- 9:00- 9:45 William Maker, Clemson University
- "Black Bodies, Black Souls: An American Narrative."
- Session 5: Race, Place, and Embodiment Moderator:
- Session 6: Identity and Race Moderator:
- Session 7: KEYNOTE: George Yancy, Emory University
- at McKissick Theatre Introduction: TBA
- 7:30- 8:00 Transportation from McKissick Theatre to the Outdoor Lab
- Session 8: Racialized Bodies Moderator:
- Session 9: Closing Session
- 3:30-4:30 Concluding remarks and open discussion
11:15-12:00 Franco Manni, King’s College London:
"Cultural Traumas, Intellectual Fallacies and Moral Mistakes’"
12:00-12:45 Joshua Sturman, Duquesne University:
"Obscuring Humanity: A Theory of Structural Racism"
12:45-2:00 Lunch Break Also, available during your lunch break: pontoon boat, canoes, high ropes, or other group initiatives.
2:00- 2:45 Miranda Young, The New School for Social Research
"Frantz Fanon’s Poetic Rupture"
2:45-3:30 Mallessa James, Goddard College
"Embodiment and Race"
3:30-4:15 Lisa M. Madura, Vanderbilt University
"Anti-Social Habit and Critical Disruption"
4:20-4:50 Transportation from the Outdoor Lab to McKissick Theatre @ Clemson University
5:00–5:30 Reception at McKissick Theatre
Introduction: Edyta Kuzian, Clemson University
7-10- 7:50 Transportation from McKissick Theatre to the Outdoor Lab
8:00-10:00 Dinner Reception and Campfire Area at the Outdoor Lab
Music Performance by Elizabeth Adams and Her All Stars
FRIDAY, April 12th
All Friday sessions, apart from the keynote address at 6pm, will all be Bowfin (Outdoor lab). The keynote address is going to take place at McKissick Theatre, 720 McMillan Rd, Clemson, SC 29631
7:30-8-30 Yoga and Meditation, Renee Gahan
7:30-9:00 Breakfast
9:45-10:30 Jyothis James, Texas A&M University
"Dialectical Stagnation and the Permanence of Racism"
10:30-10:45 Coffee Break
10:45-11:30 Nathifa Greene, Gettysburg College
"Race Talk and Land Distribution in Trinidad After Caroni (1975) Ltd."
11:30-12:15 Shaeeda Mensah, Temple University
"Fear and Black Feminine Embodiment: Black Women as Threats to the American
Populace"
12:15- 1:00 Rónké A. Òké, West Chester University
"Race Doesn’t Really Work Here": Thinking Nigeria Within the Logic of
"Racelessness" in Chimamanda Ngozi Adichie’s Americanah"
1:00-3:00 Lunch Break Also, available during your lunch break: pontoon boat, canoes, high ropes, or other group initiatives
3:00-3:45 Roxanne Burton, The University of the West Indies
"Why Race is a Crucial Dimension of Personal Identity "
3:45 - 4:30 Maia Wellborn, Fordham University
"Affective Exploitation and Black Structural Oppression"
4:35 - 5:00 Transportation from the Outdoor Lab to McKissick Theatre @ Clemson University
5:00-6:00 Reception at McKissick Theatre
6:00- 7:30
8:00-10:00 Dinner Reception and Campfire Area at the Outdoor Lab
Music Performance by Elizabeth Adams and Her All Stars
SATURDAY April 13th
All Saturday sessions will all be at Bowfin (Outdoor lab)
7:30-8-30 Yoga and Meditation Renee Gahan
8:00 -9:00 Breakfast Room
9:00- 9:30 Chiodera "Chi-Chi" Drayton-Smith, Armoni Dunwoody, Robert Magwood
9:30 – 10:30 Malik Curry, Keshauna Goines, Dajonia Jackson
10:30-10:45 Coffee Break
10:45 – 11:45 Barbara Hamberg, Racquel "Rocky" Collier, Sylvia Wu, True Liles
1:00-3:00 Lunch Break, Also available during your lunch break: pontoon boat, canoes, high ropes, or other group initiatives
4:30- 6:00 Tour of Clemson University, Clemson Area African American Museum
6:00-7:30 Dinner
Funding for the Embodiment and Race Conference, Clemson University, April 11-13, 2019 has been graciously provided by the
1) Department of Philosophy & Religion (Richard Amesbury, Ph.D.)
2) Provost Office (Carla Bennett, College Financial Officer)
3) Clemson University, College of Architecture, Arts, and Humanities (Dean Richard E. Goodstein, Ph.D.)
4) Women's Leadership (Diane Perpich, Ph.D.)
5) Pan-African Studies (Abel Bartley, Ph.D.)
6) Chief Diversity Officer (Lee Gill, J.D.)
7) Humanities Hub (Lee Morrissey, Ph.D.)
8) Department of English (Susanna Ashton, Ph.D.)
9) Creative Inquiry
10) The City of Clemson
11) Department of Sociology, Anthropology and Criminal Justice (Katherine Weisensee, Ph.D.)
12) GANTT Center (Kendra Stewart-Tillman, Ph.D.)
Cultural
Traumas, Intellectual Fallacies and Moral Mistakes
The current debate about the existence of human races
by Franco Manni
Introduction
What's the situation today? If you browse the internet by Google Chrome in
English writing these two words, “human races”, you will see that all
the entries maintain the human races do not exist. In fact, it happens
today that a relatively recent but very widespread academic fashion claims
that races do not exist, and that maintaining that they exist is a racist
attitude and behaviour.
Living beings are divided into groups. I remind you of the example of us
humans: kingdom: animals, phylum: vertebrate, class: mammals, order:
primates, family: hominids, genus: homo, species homo sapiens. Species is
the most important grouping, because it implies interfertility. Race is
synonymous with subspecies; if there is no interfertility between one
species and another, there is instead between a race (subspecies) and
another one as seen in the equine, canine and human races.
Now, a short theory of groups/sets. What is a group? It is a set (class,
category) of all those individuals who share some common characteristic
For example I can group M. L. King, Condoleezza Rice, Jussie Smollet into
the same set, because all these ‘items’ share blackness.
The divisions in groups of humanity are evident on the basis of religious
profession (Islam, Buddhism, etc.), with respect to the nationality i.e.
to language (Hispanic, Arabs, Anglo-Saxons, Chinese, etc.). However, a
division with respect to biological characteristics is also evident (blacks,
Asiatic, mulattos, whites, etc.) These biological characteristics (due to
the chromosomes) are precisely the subspecies or races of our species homo
sapiens. What characteristics, actually? They are not those of the
internal organs such as pancreas or kidneys’ shape, etc., but the
external, visible, which strike at first sight: skin colour, shape of the
nose, hairless or hairy skin, height, etc.
To my knowledge, only one biologist criticises this new theory and
restated firmly that human races do exist: Richard Dawkins in his 2004
book The Ancestor’s Tale, and in the YouTube videos he released later
on.
Cultural
Traumas
Trauma means breaking (usually of a bone). There are physical traumas,
such as a lethal epidemic disease, a tsunami, a famine, and cultural
traumas, i.e. wicked human deeds which wound the stability, the security,
the individual life in a society and warp the thoughts, attitudes,
feelings of many people in that society. Such are slavery, political
tyranny, military conquests, persecution.
Two major cultural traumas in recent Modern history have been: 1) the
slave-trade and slave-exploitation of black African people in the Americas
till the 19th century, and 2) the persecution and genocide of the European
Jews in the 1930s and 1940s.
Now, in both case the key word used to describe the oppressed categories
was ‘race’. In the first event the word is correct. In the second it
was not because the European Jews did not belong to a particular race, to
any alleged (but inexistent) Jewish race.
Anyway, in both cases the universally widespread idea was that the two
categories (the African blacks and the Jews) belonged to a race, i.e. to a
biological group.
But, why were those two races oppressed? Here racism comes out. And: what
is racism? Racism means to believe that one race is superior to the others
and that, because of this alleged superiority, it is allowed to question
and limit the rights of the other people, and oppress, exploit and
persecute them. In the first case whites thought that Caucasian race was
superior and the black inferior, in the second case the Nazis thought that
the ‘Aryan’ race (North European white with blond hair, pale skin and
blue eyes) was superior and the Jewish one both inferior and dangerous for
the development of human civilisation.
These two traumas affected not just the oppressed blacks and Jews , but
also the European persecutors, some of whom now feel a sense of guilt. I
think that these traumas are an important cause (even if not the only one
) which now makes people confuse the very negative ideology of racism -
which is a political fact - with the existence of races, which is only a
biological fact.
This phenomenon is one of the big three ones of Western civilisation.
In 17th-18th centuries there was the idea that a religion, be it Catholic
or Protestant, was superior to the other one, and that, because of this
alleged superiority, they had the right to oppress and persecute the
others. In the 19th century there was the idea that a nation was superior
and therefore entitled to oppress and rule the others. In this case the
German nation towards the Italian, Slovenian, Bohemian, Croatian, Polish
and French nations. And the Russian nation towards the Polish, Romanian,
Slovak, Ukrainian, Armenian, Turkish, Finnish nations. And the Jewish
nation (Zionism) superior to the Arab nation (Palestinians). And, more in
general, the Europeans thought their nationalities were superior to the
Asian ones, the Indians, the Chinese and the like.
Now, to communicate and show you the mistake of confusing race with racism,
I examine the other two phenomena. Do religions exist? Does exist
religious fanaticism i.e. the idea that one religion is superior to others
and because of this alleged superiority it is entitled to oppress the
others? If you answered YES to these two questions, now I ask you the
final one: since we dislike this religious fanaticism, does it imply that
different religions do not exist?
As for nationality, do different nations, that is peoples, exist, such as
Hispanic, Chinese, Arabs, Kurds, Germans? Does exist “nationalism”,
i.e. the theory that claims that one nation is superior to the others? If
you answered YES to these two questions, now I ask you the final one:
since we dislike nationalism, does it imply that different nations do not
exist?
The case of racism has been longer and more wicked than the other two.
Because it relies on races, a biological entity and not a cultural entity,
unlike religion and nationality, it is something which cannot be changed.
When a powerful state conquers other peoples, it exports its nationality,
i.e. language and economic-sexual traditions, as it happened with the
Spanish conquering Mexico and the British conquering Australia. A religion
can spread by several means, not just by wars of conquest, as happened
with Christianity in the Ancient Roman Empire or Buddhism in the Ancient
Chinese Empire. In both cases, therefore, you can achieve changes: you can
teach a language to a person and convert him to another religion. Whereas,
you cannot change his race.
This was the tragic case of the African slaves, that even if they learned
English, and had tea and porridge, embraced Christianity, disposed
polygamy and were dressing like the Europeans, all the same they remained
black in their very physical bodily skin.
Throughout the 20th century the former slaves (and the former colonised
peoples of Africa) acquired more and more rights from their former masters
and rulers, so that more and more in all the jobs (even president of the
US!) the black have become more and more similar for wealth, education and
power to the white. It is like a law of human nature: the more you improve
after a trauma, the more you feel the pain. I fall down the stairs but in
the first minutes I do not feel pain. A child is abused by his parents is
not aware of the destruction of his mind while he is small and the abuse
is going on. When he grows up and recovers, it is then that he intensely
feels pain and sorrow.
I think that this is the historical psycho-social context for these facts.
The black became more and more equal in social position. Paradoxically the
more they became equal the more they complained about inequality. And the
more (this factor is more important) the whites were ashamed of past nasty
inequality.
In fact, the essay of biologist Richard Lewontin where the existence of
races is questioned was written already in 1972. But it took decades to
make its way through the scholarship and eventually to the public. Still
in the edition of 1997 the Encyclopaedia Britannica read:
"Naturally occurring (i.e., produced by natural, usually geographic
separation of human groups) R A C E S of the human species are by no means
identical in number of members or degree of genetic differentiation. There
are small groupings of a few hundred to a few thousand individuals, some
slightly and only recently isolated reproductively from adjacent people.
It is both useful and meaningful to identify the very large human
groupings that often correspond to continents or other major geographic
areas as geographic R A C E S, a term extensively used with other life
forms”.1
But a few years later something happened: in 2001 the most popular science
magazine in UK, Nature, came with a CD containing the core statement of
the old article by Lewontin :
“]in 1972 Richard Lewontin of Harvard University] ‘‘found that nearly 85 per cent of humanity’s genetic diversity occurs among individuals within a single population. /…/In other words, two individuals are different because they are individuals, not because they belong to different races”.2
And in 2002 on of the most important US science magazines, The New Scientist, reported the same Lewontin’s statements. And quickly this old idea became fashionable and spread among the biologists first, and then the learned public. So that in 2003 A.W.F. Edwards, an old and renowned biologist, fellow of the Royal Society, wrote an article on the academic journal BioEssays which debunked the theory held by Lewontin. We will see this debunking later on, in the words of Richard Dawkins. Here I notice that the confutation made by Edwards was not successful and did not prevent the Lewontin’s verbum from spreading more and more.
What
events (cultural, social, political) triggered this resurrection of
Lewontin’s essay from the dead cemetery of forgotten scholarship? What
cultural dynamic brought the Western academia after the end of communism
in 1989 and just before 9/11 tragedy and its entailed islamophobia, to an
unleashed extreme liberalism? It would be worth investigating! A good
project for a historian of ideas!
Let us read the 2019 Edition of the same Encyclopaedia Britannica which
says:
“Genetic studies in the late 20th century refuted the existence of
biogenetically distinct races, and scholars now argue that “races” are
cultural interventions reflecting specific attitudes and beliefs that were
imposed on different populations in the wake of western European conquests
beginning in the 15th century.”3
Very
different from the previous edition, isn’t it?
Intellectual
Fallacies
Let us examine two intellectual fallacies.
Apart from races, there are other hierarchies where we see a first
intellectual fallacy. This fallacy is confusing the questionable
hierarchical framework with the different qualities between groups: active/contemplative
life, celibate/married, male/female.
The last one about gender is current and hot today: in order to fight
hierarchy of superior / inferior, more and more people pretend that there
are not differences between man and woman. Here we see the same the idea
of deleting the differences. So that, this pretension creates injustices.
An iconic example is that transgender woman (biologically a man) Rachel
McKinnon who won the 2018 World Championship of cycling . (see Image1)
Image 1
The
second fallacy is Lewontin’s argument itself, which claims that there
are not biological group differences within human beings, because the
differences between individuals are bigger than those one between groups.
Richard Dawkins, in his book The Ancestors’ Tale, explains why this
argument is false:
“It is genuinely true that, if you measure the total variation in the
human species and then partition it into a between-race component and a
within-race component, the between-race component is a very small fraction
of the total. Only a small admixture of extra variation distinguishes
races from each other. That is all correct. What is not correct is the
inference that race is therefore a meaningless concept. /…/However small
the racial partition of total variation may be, if such racial
characteristics as there are highly correlated with other racial
characteristics, they are by definition informative, and therefore of
taxonomic significance”.4
Let us pick up an example: the poster of UK Government campaigning against
prostate cancer for BLACK men. (see Image 2) The probability that a black
man suffers from a prostate cancer is double than for white men. What is
the meaning of BLACK ? A group. Ok. What group actually? The Muslims? The
citizen of Nigeria? The Arabic speaking people? I mean, is the criterion
for grouping religious, or juridical or linguistic? No, it is biological.
A grouping of homo sapiens in sub-groups in respect of the species homo
sapiens by a biological criterion. What are these subgroups of species
homo sapiens? Sub-species. What is the word for subspecies suitable to
laypeople? Races.
Image 2
Let us
listen to what the best evolutionary biologist of the 20th century,
Stephen J. Gould, says :
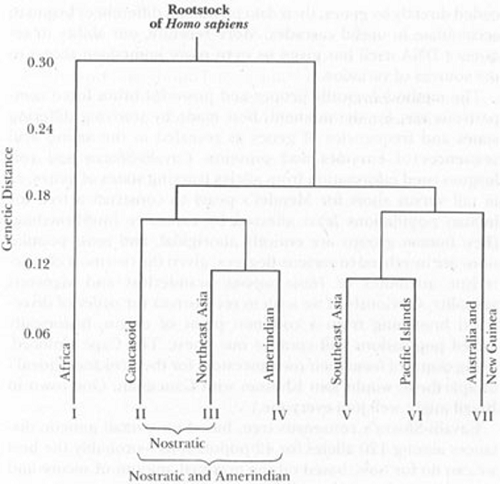
“Luca Cavalli-Sforza’s consensus tree, based on overall genetic
distances among 120 alleles for 42 populations—probably the best we can
do for now, based on the maximal amount of secure and consistent
information—divides modern humans into seven major groups, as shown in
the accompanying chart.
These seven basic groups, established solely on genetic distances, make
excellent sense when we consider the geographic distribution of Homo
sapiens. Humans presumably evolved in Africa, and the first great split
separates Africans from all other groups—representing the initial
migration of some Homo sapiens out of the mother continent. The next split
separates the coherent region of the Pacific and Southeast Asia from the
rest of the world., etc, etc.”.5
Now I ask
you, using the very words of Stephen J. Gould: what are these “seven
basic groups established solely on genetic distances”? (see Image 3).
Are they nationalities? Are they Religions? Are they political ideologies?
Or, instead are races?
There are, sadly, walls that prevent communication between the specialised
scientific disciplines. Mental walls, I mean. For instance, most of the
evolutionary biologists know the studies of grouping the member of species
homo sapiens in seven major groups. But most of the biologists affirm the
races do not exist. It is as if the mind was split in two
non-communicating vessels.
Image 3
Moral
Mistakes
The cultural traumas and the intellectual fallacies trigger moral mistakes.
1) First moral mistake. Those who deny the existence of races (and
therefore of these different characteristics of grouping), certainly will
never be able to convince ordinary people to whose eyes these differences
are evident and will never disappear. Therefore, first of all, the
supporters of this new theory contribute to a very destructive
anti-intellectual common place, i.e.: what the academia says is something
esoteric and arcane without any connections with reality, so it is
worthless to develop knowledge and scholarship.
As Dawkins says:
”It is a point I heard being amusingly lampooned by a West African
medical researcher at a gathering of about 20 scientists. At the beginning
of the conference, the chairman asked each of us around the table to
introduce ourselves. The African, who was the only black person there —
and he really was black, unlike many 'African-Americans' — happened to
be wearing a red tie. He finished his self-introduction by laughingly
saying, 'You can easily remember me. I am the one with the red tie.' He
was genially mocking the way people bend over backwards to pretend not to
notice racial differences.”.6
2) Second
moral mistake. They also commit the grave error, both intellectual and
moral, to give this message to people: in order to affirm that all men
have equal dignity and therefore must have equal rights, we are obliged to
state that they are equal in their material and de facto situations.
But this is a serious moral mistake. Oure equal dignity should be
recognized not because we are equal in our material, social and cultural
conditions (such as, all equally Christians, all equally heterosexuals,
all equally healthy, all equally communist/fascist, all of the same
biological group) !
On the
contrary! Equal dignity must be recognized precisely when we see that
materially, economically, linguistically by political opinion, by
religious faith, by somatic characteristics, by sexual orientation we are
different. The important human value at stake here is: equal moral and
legal dignity, despite the material and social differences!
Otherwise, the same concept of tolerance disappears. In fact. it is
illogical to say that you practice the virtue of tolerance towards those
who are equal to you by religion, social class, nationality, etc. In fact,
it is possible to be tolerant only towards those who are different from
you!
3) Third
moral mistake. At the end of his critique of Lewontin’s thesis, Edwards
says:: “it is a dangerous mistake to premise the moral equality of human
beings on biological similarity because dissimilarity, once revealed, then
becomes an argument for moral inequality.” Since the ‘revelation’,
of these differences is always at hand, easy and powerful, if the ‘revelation’
entails again inequality and hierarchy, well, we will have more inequality!
It is as if I said: “There are no differences between rich and poor ,
doctors and nurses, MPs and cleaners,” and I said that in order to
defend their equal dignity before the state, the law, the society, well,
whenever the differences between them had been clearly and overwhelmingly
revealed, then we would think that those groups do not share the same
human dignity.
If you really do not want to hit the hypersensitivity of those people who
deny the existence of the races, you could use another word or phrase (biological
groups, subspecies, genetic classes ......). Ok. However, in any case, to
deny the reality is an intellectual and moral mistake.
This tendency, by which even scientists accommodate their views to the
fashionable social ideologies of the moment, is very dangerous. Before
WWII many biologists ignored lots of contrasting empirical data in order
to maintain that the biological difference between races entails
psychological, intellectual, moral and political characteristics. The
outcome was the creation of racism. Today most biologists, against lots of
empirical evidence, maintain that races (genetic group differences) do not
exist and are creating a dystopic totalitarian world were in order to be
equally treated by the state and the society we need to deny the existence
of differences among us.
Bibliography
Stephen J. Gould, Bully for Brontosaurus: Reflections in Natural History
(New York: Norton, 1991).
A.W.F. Edwards, ‘Human genetic diversity: Lewontin’s fallacy’,
BioEssays 25:798–801, 2003.
Richard Dawkins, The Ancestor’s Tale, (London: W&N, 2004).
Richard Lewontin, ‘The apportionment of human diversity’, in
Dobzhansky T, Hecht MK, Steere WC, editors. Evolutionary Biology 6 (New
York: Appleton-Century-Crofts, 1972), pp 381–398.
Encyclopaedia Britannica 1997 edition.
Encyclopaedia Britannica 2019 edition.
‘The Human Genome’, Nature 2001;409.
Anil Ananthaswamy ‘Under the skin’, New Scientist 2002;174.
Franco Manni indice degli scritti
Maurilio Lovatti main list of online papers